|
||||||
The
Creation Explanation
|
|
 |
The Primeval World -- Fossils, Geology & Earth History |
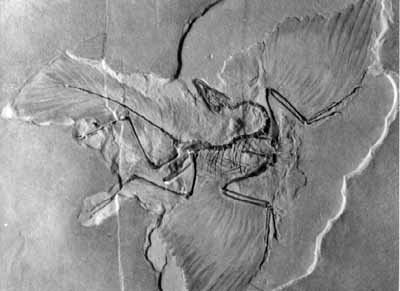
Do Fossils Speak For Evolution Or For Creation? As we begin to develop the subject of the fossil record we must give credit to Dr. Duane Gish for some of the basic elements of our approach which he first introduced in his book, Evolution: the Fossils Say NO1 and the more recent Evolution: the Challenge of the Fossil Record.2 First, we will compare two contrasting models designed to explain the evidence bearing on the origin of living creatures, both past and present. These two models make possible two entirely different predictions concerning what we might expect to find in the fossil record: 1. If evolution is the correct explanation for the origin of the fossils, finely graded sequences of intermediate fossil forms should be found to be systematic in the fossil record. Such sequences of intermediate types should be found connecting all kinds of creatures, living and extinct, leading back to some original simple, single celled organisms. Some intermediate fossils might be forever lost through geological processes, but in general they should be found. 2. If creation is the correct explanation of beginnings, the fossil record should reveal a systematic absence of intermediate types which might connect the different categories of plants and animals. Let us now consider the fossil record to see which prediction more satisfactorily fits the actual evidence. 1. The supposedly earliest or deepest lying sedimentary rocks containing unquestionable fossils of multicellular life forms are called Cambrian rocks, named after Cambria in the southern British Isles. Dated as being up to 600 million years old, Cambrian rocks have yielded fossil evidence for 2,000 (some say 5,000) species representing every major phylum of animal life, including the vertebrate sub-phylum.3 2. The allegedly much older Precambrian rocks are devoid of fossils, other than microscopic trace fossils of single-celled algae and bacteria. So where are the ancestors of the rich and varied assortment of Cambrian fossils? It seems apparent that three-quarters of all evolution had to take place before the Cambrian rocks were deposited, but where is the evidence? Instead of fossil evidence for a gradual process of evolution, we find a massive unfilled gap between all of the major phyla of complex life forms and the single-celled life forms from which they supposedly evolved. This is the first great gap in the fossil record which provides evidence agreeing better with the concept of creation rather than with that of evolution.4 Charles Darwin himself admitted that the gaps in the fossil record supplied perhaps the most damaging evidence against his theory.5 3. Vertebrate fish supposedly evolved from invertebrates, yet in his authoritative book, Vertebrate Paleontology, Harvard paleontologist Alfred Romer stated that the fossil gap between the vertebrate fish and their unknown invertebrate ancestors is 100 million years.6 Furthermore, the various orders of fish appear from the fossil record to have arrived on the scene completely separate and distinct from the beginning.7 The theories about which invertebrate creature gave rise to the vertebrates are almost as numerous as are books on paleontology. 4. Amphibians are assumed to have evolved from fish, yet there is no connecting sequence of intermediate fossilized forms. There should have been a multitude of intermediate forms leading from the fin of the crossopterygian fish to the leg of the of the ichtheostegid amphibian. None has been found.8 Another difficulty stems from the forms of the vertebrae in supposed evolutionary series. Both the crossopterygian fish and the ichthyostegid amphibians had arch type vertebrae.9 On the other hand, three allegedly more modern fossil orders of amphibians which are assumed to have evolved from them have supposedly more "primitive" vertebrae of the so-called "husk vertebrae" type. Strangely, the three living orders of amphibians also have the "primitive" type of vertebrae. Finally, none of groups of amphibians is connected to an ancestral group of fish by a series of intermediate types.10 5. We are told that reptiles evolved from amphibians. While it is true that fossil amphibian skeletons rather similar to reptile skeletons have been found, serious unanswered questions remain for the evolutionary view. A gap between the amphibians and the reptiles is found in the fact that the foremost candidates for the key transitional amphibians, Seymouria and Diadectes, are found in the wrong rocks. They reportedly appeared some twenty million years after the appearance of the original reptiles and also after the appearance of the reptile group from which the mammals are said to have evolved.11 How could the parents appear after the children, so to speak? Another problem is the requirement that the simple gelatinous amphibian egg, designed to develop in water, be transformed by slow, minute changes into the complex amniotic egg of the reptiles, designed to incubate in the air. There is no direct fossil evidence for this transformation, and it is difficult indeed to imagine how it would have occurred.12 6. Mammals are assumed to have evolved from the reptiles, but once again there is an absence of needed transitional sequences. A key issue is structural differences in the jaws and ears of reptiles and mammals. Secular paleontologists and anatomists have long studied the fossil materials and believe they have explained how a gradual transition of jaw bones and inner ear bones from the reptilian to the mammalian condition occurred. However, crucial fossil material is missing so that the case for reptile-to-mammal evolution still rests on circumstantial evidence. Dr. Duane Gish has discussed this problem in detail.13 All reptiles have four bones on both sides of the lower jawbone, whereas all mammals have a single bone, the dentary. The mammalian dentary forms a joint with the squamosal bone of the skull, whereas in reptiles the joint is between the articular bone and quadrate bone of the skull. Several "mammal-like" fossil reptiles, notably Diarthrognathus, Morganucodon and Kuehneotherium, are said to have had both types of jaw joints at the same time. Thus, they were supposedly reptiles on the way to becoming mammals. But no skulls have been found which show the dentary actually in contact with the squamosal bone. Furthermore, all of these fossil reptiles have a powerful reptilian jaw joint. Three of the reptilian lower jaw bones are supposed to have migrated into the ear to become the three delicate, precisely engineered inner ear bones, or ossicles, of the mammals. However, all fossil and living mammals have three ossicles, but all fossil and living reptiles have a single inner ear bone. And it is difficult to imagine how jaw bones, by chance mutations and natural selection, could have evolved into the marvelous inner bones of mammals. One detailed evolutionary study of the fossil materials and the pertinent physiology concluded that the transition took place, but the evolutionary argument still involves many assumptions.14 Some appeal to evidence from the embryonic development of certain mammals, for example kangaroos, during which the embryonic bones are observed to make shifts which correspond to the proposed historical sequence of adult forms of jaw and ear bones. The required intermediate forms must be adult forms. Another problem is the origin of the exceedingly complex organ of Corti in the mammalian ear. The high technology by which the organ of Corti translates sound vibrations into nerve impulses transmitted to the brain is still only partially understood. No reptile possesses this structure, and there is no fossil evidence to prove mammals obtained it by evolution. The mammalian inner ear is "one of the most remarkable displays of precision microengineering in the vertebrate body."15 Without historical evidence for its origin by evolution, determined faith is required to see the organ of Corti happening without God the Creator. No testable scientific theory exists to explain the evolution of the design of the organ of Corti, nor is the location of the design information in the genes known or how that information is used to produce the marvelous structure. In the presumed evolution from reptiles to mammals other amazing new designs had to be produced by chance, including new modes of reproduction and breathing, mammary glands, temperature regulation and hair. In reptiles the thorax is not expansible as in mammals, nor do reptiles have a diaphragm. There is no fossil evidence for the evolution of any of these features of mammals from the reptiles. The imagination is strained to the breaking point to explain how evolution did it all. To believe that mammals were specially designed and created by God the Creator is not at all "unscientific," and actually seems to be a more reasonable faith. 7. The fossil gap between the thecodont reptiles and modern birds is said to be a period of about 80 million years. For years there was only the fossil Archaeopteryx lithographica to offer as an intermediate species.16 This fossil bird does not, however, display characters partially transformed from the reptilian to the avian(bird-like) type. Stephen J. Gould of Harvard University and Niles Eldridge of the American Museum of Natural History have pointed out that Archaeopteryx is actually a "mosaic" of fully reptilian and avian characters.17 For example, it was fully feathered and therefore definitely to be classed as a bird. The flight feathers of the wings appear to have been identical in structure to those of modern birds. They were asymmetric as are the flight feathers of modern strongly flying birds. However, two recent studies have led to contrary conclusions.18 Speakman and Thomson asserted that the asymmetry of the Archaeopteryx feathers was more comparable with those of modern non-flying birds. Their conclusions were contested by R. Åke Norberg, who found the asymmetry of the fossil bird's flight feathers fell in the range of modern flying birds. Speakman and Thomson rebutted Norberg and added that the structure of the elbow joint of the wings and the estimated strength of the wing muscles suggest that Archaeopteryx was a non-flying bird. Numerous intermediate forms would surely have been required gradually to transform reptile scales into bird feathers and change reptiles into birds, but fossil evidence for this process of change is missing. The fine structure of flight feathers is intricate and highly contrived. Without the historical evidence that only fossils can provide, the idea that scales became feathers continues to be an article of faith. Some of the reptilian features of Archaeopteryx are possessed by some modern birds, and others are not found in some modern reptiles. Archaeopteryx had teeth in the bill, claws on the wings, an unfused backbone, a long bony tail, and a breastbone without a keel. The sum of the evidence does not qualify this as an intermediate type.19 Especially is this so in view of the recent report of a femur and other bird bones of more modern type found in Jurassic rocks reputedly of the same age as those in which Archaeopteryx is found.20 Another problem with the concept of bird evolution is the fact that reptile lungs are constructed of millions of tiny air sacs with in-and-out airflow, as are mammal lungs. But bird lungs have tubes rather than sacs, with air flowing through in one direction only.21 In addition to this totally different structure and operation of the lungs, birds must have a clever system of flexible air sacs and channels to collect air and conduct it to the lungs for each breath. How could a creature with a lung made half of sacs and half of one-way tubes breathe and survive? The creation view is that this bird and all others were created with their basic characters complete and fully functional. The same may be said for the reptiles. There is no evidence to refute this view.
figure 3-1. The Archaeopteryx bird fossil shows fully developed feathers on wings, body, and tail. It was definitely a bird, although possessing a number of reptilian characteristics. 8. This brings us to the question of the origin of flight in general. In no case is there fossil evidence for the origin of flight, not in the case of birds, mammals, reptiles, or insects. The origin of flight is a complete mystery as far as fossil evidence is concerned. The flying insects were always flying insects,22 the now extinct flying reptiles have no transitional fossils connecting them to non-flying reptiles,23 and the flying mammals (bats) were always well-engineered bats.24 The evolutionary explanations devised thus far can seem credible only to true believers.25 Flying creatures were apparently designed to fly from the very beginning, just as reported in Genesis. 9. The absence of connecting links is especially noticeable in the fossil record of the more peculiar kinds of animals. For example, the cetacea (whales, dolphins and porpoises), sirenia (manatee, dugong and sea cows), pinnipedia (sea lions, seals, and walruses), turtles, kangaroos, bats, dragon flies, spiders, and other types are isolated in the fossil record. In each such group the fossils are all distinguished by the peculiar features necessary for their separate classification. Their evolutionary origins are represented by missing links and speculations, portrayed in standard reference books by dotted lines in the charts showing supposed evolutionary descent. In the case of whales, recent fossil finds have provided some reasonable intermediate forms which can fit into the big gap between the fossil mesonychid land mammals and extinct fossil whales. Stephen Jay Gould asserted in one of his regular monthly essays in Natural History published by the American Museum of Natural History that these new fossil finds "confirm the theory of the whale's return to the sea."26 At about the same time, however, Michael J. Novacek of the Museum research staff, reviewing these fossils in Nature, admitted that, "Ambulocetus, Rodhecetus and other more aquatically specialized archaeocetes cannot be strung in procession from ancestor to descendant in a scala naturae(ladder of nature)."27 There is no question that these fossils are suitable candidates to be fit into possible sequences leading from land mammal to whale. However, very substantial gaps remain, so that incontrovertible proof for the evolutionary origin of whales has yet to be adduced. The case for the evolution of turtles has also been advanced by recent studies of similarities between extinct turtles and a group of extinct reptiles, the pareiasaurs. Michael Lee proposes that the pareiasaurs evolved a number of turtle characters before Proganocheles, the most ancient turtle arrived on the scene. Therefore, he proposes the pareiasaurs as the best candidates for the ancestors of the turtles. The reasoning based on detailed studies of the fossil materials sounds persuasive. Lee does admit, however, that there is a large gap between Proganocheles and the pareiasaurs:28 But where do turtles come from? The oldest fossil turtles (along with the earliest dinosaurs) appear abruptly in the Triassic rocks, fully developed and without any obvious precursors. ...Proganocheles retained many features inherited from its pareiasaurian forebears. ...Nevertheless, a forty-million-year gap, spanning almost the entire Triassic still exists between the last pareiasaurs and the earliest-known turtles. When turtles first appear in the fossil record, in the late Triassic, they are represented by at least four distinct lineages, suggesting that the group evolved and radiated slightly earlier. Once again we see that the crucial evidence for an evolutionary origin is missing from the fossil record. It is not true, as some creationists have asserted, that there is no evidence for evolution. There is a great deal of evidence for evolution, but there is also much evidence for intelligent, purposeful creation. In both cases the evidence is circumstantial, and its interpretation depends upon philosophical presuppositions. It is our position that the scientific case for evolution has not been proved unless the fossil evidence is found that documents construction by gradual evolutionary processes of new complex structures and organs. Forty million years dreaming up and constructing turtles is a big gap in the required evidence. 10. Perhaps the best known demonstration of an evolutionary scenario is that of the horse series displayed in school and college textbooks and in museums. These charts and displays make the theory of horse evolution very neat, seemingly historical, all cut-and-dried. Actually there are important problems with the theory and some serious disagreement, even among evolutionary scientists.29
To summarize, the alleged horse evolution series actually appears to be three groups of genera. The first in the series has no connection by fossil intermediates to the supposed ancestors. The three groups may well have no connection one with the other, and the overall fossil horse data can be fitted into the framework of the biblical creation model. The three groups of genera may represent three created kinds which should be fitted into the classification system as three separate "families" of ungulates. There is no need to assume that horses were evolved rather than created. The faith of atheistic materialism leads one to evolved horses. The faith of biblical theism leads to created horses. 11. What evidence exists to prove that man evolved from a common ancestor with the apes? The history of research into reputed human evolution is said by some leading secular scientists to be a blot on the reputation of science. As we shall see in Chapter 5 there is no empirical evidence to prove the secular case. What the evidence does prove is that interpretations and conclusions are strongly dependent upon philosophical commitments, personal ambition and monetary considerations.
References 1. Gish, Duane, Evolution, the Fossils Say NO (Creation-Life Pub., San Diego, 1973). 2. Gish, Duane, Evolution: The Challenge of the Fossil Record (Master Books Pub., San Diego, 1985). 3. Simpson, G.G., The Meaning of Evolution (Bantam Books, New York, 1971), pp. 16-19; Axelrod, Daniel I., Science, Vol. 128, 4 July 1958, p. 7. 4. Newell, Norman D., Proc. of the Philosophical Soc., Vol. 103, April 1959. 5. Darwin, Charles, The Origin of Species (J.M. Dent, London, 1971), pp. 314-316. 6. Romer, Alfred S., Vertebrate Paleontology, 3rd Edition (Univ. of Chicago Press, 1966), pp. 23, 318. 7. _________, ibid., pp. 25, 47; Harland, W.B., et al., Editors, The Fossil Record (Geological Soc. of London, 1967), p. 630. 8. Romer, Alfred S., ibid., pp. 79, 318; Anderson, J. Kerby, and Harold G. Coffin, Fossils in Focus (Zondervan, Grand Rapids, MI, 1977), pp. 51-53. 9. Romer, Alfred S., ref. 6, pp. 79, 87, 88. 10. Ibid., pp. 87, 98, 100, 101; Gish, Duane, ref. 1, pp. 74-76. 11. Harland, W.B., et al., ref. 7, pp. 686, 696; Gish, Duane, ref. 2, p. 77; Romer, Alfred, ref. 6, p. 173. 12. Romer, Alfred S., ref. 6, pp. 95, 102, 103. 13. Gish, Duane, ref. 2, pp. 88-103. 14. Crompton, A.W. and Parker, Pamela, American Scientist, Vol. 66, March-April 1978, pp. 192-201; Gish, Duane, ref. 2, p. 58; Romer, Alfred S., ref. 6, pp. 104, 187-188, 191; McGraw-Hill Encyc. of Science and Technology (New York, 1971), Vol. 4, pp. 359-360. 15. Swanson, G.J., M. Howard, and J. Lewis, Journal of Developmental Biology, Vol. 137, 1990, pp. 243-257, quoted by Brigid Hogan and Christopher Wright, Nature, Vol. 355, 6 Feb. 1992, p. 495. 16. Romer, Alfred S., ref. 6, pp. 13, 166-167, 368, 374. 17. Gould, S.J. and Niles Eldridge, Paleobiology, Vol. 3, 1972, p. 147. 18. Speakman, J.R. and S.C. Thomson, Nature, Vol. 370, 18 Aug. 1994, pp. 514; Norberg, R. Å, Nature, Vol. 374, 16 March 1995, pp. 221-222. 19. Morris, Henry M. and Gary E. Parker, What Is Creation Science? (Creation-Life Pub., San Diego, 1982), pp. 101-105. 20. Jensen, James A., reported in Science News, 14 Sept 1977, p. 19; J.L.M., Science, Vol. 199, 10 Jan. 1978, p. 284. 21. Schmidt-Nielsen, Knut, Scientific American, Vol. 225, Dec. 1971, pp. 72-79. 22. Harland, W.B., et al., ref. 7, pp. 107, 117, 508. 23. Romer, Alfred S., ref. 6, pp. 144-147. 24. Ibid., p. 338; Jepsen, G.L., Science, Vol. 154, 9 Dec. 1966, p. 1333. 25. Harland, W.B., et al., ref. 7, pp. 107, 117, 508; Romer, Alfred S., ref. 6, pp. 144-147, 338; Jepsen, G.L., ref. 24, p. 1333. 26. Gould, Stephen Jay, Natural History, Vol. 103, May 1994, p. 8. 27. Novacek, Michael J., Nature, Vol. 368, 28 April 1994, p. 807. 28. Lee, Michael, Natural History, Vol. 103, June 1994, pp. 63, 64; see also Lee, Michael S.Y., Science, Vol. 261, 24 Sept. 1993, pp. 1716-1720. 29. Simpson, G.G., Tempo and Mode in Evolution (Columbia Univ. Press, New York, 1944), p. 167; Cousins, Frank W., Creation Research Soc. Quarterly, Vol. 8, Sept. 1971, pp. 99-108; Nilsson, Heribert, Synthetische Artbuildung, (Verlag C.W.K. Gleerup, Lund, Sweden, 1953: reprint of English summary published by Evolution Protest Movement of North America, Victoria, B.S., 1953), pp. 1193-1194; Kerkut, G.A., Implications of Evolution (Pergamon Press, New York, 1960), pp. 144-149; Wentworth Baroness, Thoroughbred Racing Stock (Charles Scribners, Sons, New York, 1938), p. 379. 30. Simpson, G.G., Horses (Oxford Univ. Press, New York, 1951), pp. 105-112, 115-116. 31. Ibid., pp. 116-117; Simpson, G.G., ref. 3, p. 135. 32. Simpson, G.G., ref. 30, p. 124. Other fossil horse data cited below can be found in the same work. 33. Gish, Duane, ref. 2, pp. 83-84; Romer, Alfred, ref. 6, pp. 260-261. 34. Nevins, Stuart E., Creation Research Soc. Quarterly, Vol. 10, March 1974, p. 196. 35. Simpson, G.G., ref. 30, pp. 177-179; Davidheiser, Bolton, Creation Research Soc. Quarterly, Vol. 12, Sept. 1975, pp. 88-89. 36. Bethel, Tom, "The Taxonomic Case Against Darwin," Harper Magazine, Feb. 1985, pp. 49-61. Niles Eldredge is quoted on page 60. |
|
|
|